ПРИМЕЧАНИЯ ПО ЭМБРИОЛОГИИ
Эмбриология изучает последовательность форм развития от зиготы до организма, наделенного всеми его органами и системами.
В связи с этим стоит помнить о различии между развитием (последовательность структурных и организационных фаз с возрастающей сложностью) и ростом, прежде всего в количественном отношении.
У многоклеточных позвоночных мы наблюдаем, поднимаясь в эволюционном ряду вплоть до человека (через циклостомов, рыб, земноводных, рептилий, птиц и млекопитающих), появление взрослых форм возрастающей сложности, для которых усложняются стадии эмбрионального развития.
Вначале зигота, всегда снабженная резервным материалом, делится (последующим митозом) на 2, затем 4, затем 8 и т. Д. Клеток, называемых бластомерами, без роста, пока не будет достигнуто нормальное соотношение ядро / цитоплазма, характерное для данного вида. .
Эта начальная сегментация может следовать разным образцам в зависимости от количества и распределения дейтоплазмы.
Вначале дейтоплазма недостаточна («олиголецитарные яйца»), поэтому сегментация является тотальной и приводит к появлению самых разных бластомеров. По мере увеличения сложности эмбриона требуется больше времени и материала, прежде чем его развитие позволит ему начаться. самостоятельная жизнь. Это требует увеличения дейтоплазмы («телолецитарных яиц»), которая имеет тенденцию располагаться в части зиготы. Это вызывает возрастающую «анизотропию», которая связана с модификациями сегментации, регулируемыми двумя общими принципами:
- Закон Хертвига гласит, что при митозе ахроматическое веретено (экватор которого определяет плоскость деления дочерних клеток) имеет тенденцию располагаться в направлении наибольшей длины цитоплазмы;
- Закон Бальфура гласит, что скорость сегментации обратно пропорциональна количеству дейтоплазмы.
Таким образом, мы видим, что уже у циклостомов и у рыб сегментация неравномерна, с быстро сегментированным анимальным полюсом (который дает верхние структуры эмбриона) и желточным полюсом, который будет содержать большую часть резервного материала. Эта тенденция еще больше y анизотропный у земноводных (у которых необходимо подготовить органы, ответственные за дыхание воздуха), у которых желточный полюс, медленно сегментируясь, остается относительно инертным и в конечном итоге покрывается клетками, происходящими из быстро сегментирующего животного полюса. основные эмбриональные стадии включают: зиготу, бластомеры, морулу (скопление бластомеров, похожих на ежевику), бластулу (морула с регрессировавшими внутренними клетками), гаструлу (бластулу, в которой клетки одной стороны стали инвагинированными), в которой примитивная полость организма, с внешним клеточным слоем (эктодерма, из которой в первую очередь будет происходить нервная система) и внутренним erno (энтодерма), между которыми будет вставлен третий слой (мезодерма). Затем из этих слоев или «зародышевых пластов» в упорядоченной последовательности будут происходить все ткани, органы и системы.
У даже более развитых видов увеличение дейтоплазмы (или «теленка») таково, что ее невозможно даже сегментировать. Таким образом, мы видим, что у птиц сегментация затрагивает только тонкий поверхностный диск, что приводит к «дискобластуле» и серии дефектов. явления, которые гарантируют формирование эмбриона иным способом, чем упомянутый выше.
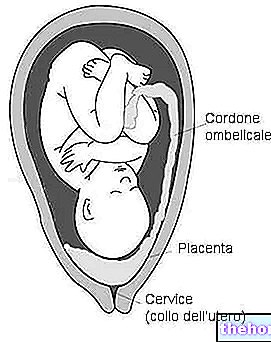
Дальнейшее увеличение дейтоплазмы, вероятно, не было бы более эффективным, поэтому у млекопитающих развитие и рост до способности к независимой жизни достигается с помощью другой системы. Фактически, у млекопитающих мы отмечаем, что дейтоплазма используется только на самых первых стадиях. развития; затем эмбрион устанавливает метаболические отношения с материнским организмом (с появлением плаценты) и больше не использует дейтоплазму, избыток которой устраняется. до тотального (и, следовательно, на ранних стадиях он подобен таковому у «амфиоксуса»), но после морулы эмбриогенез продолжается в соответствии с наиболее развитым образцом птиц, с «бластоцистой», за которой следует имплантация на стенке матки, так что метаболизм эмбриона обеспечивается материнским организмом (через плаценту), а не дейтоплазмой.
ДИФФЕРЕНЦИАЦИЯ ЭМБРИОНОВ
Когда сегментация зиготы довела соотношение ядро / цитоплазма до нормы вида, рост должен начаться параллельно с развитием.По этой причине метаболизм начинается с появления ядрышек и синтеза белка. Инициируемый таким образом синтез белка происходит благодаря генам, ответственным за ранние стадии эмбрионального развития. Эти гены подавляются веществами, присутствующими в различных бластомерах животного и теленка. В свою очередь, продукты этих ранних генов могут подавлять опероны генов, отвечающих за более поздние стадии. Продукты этой второй серии генов будут способны действовать как в смысле создания новых эмбриональных структур, так и в смысле подавления предыдущих оперонов и подавления следующих оперонов в упорядоченной последовательности, которая приводит к созданию нового организма. , благодаря генетической информации, накопленной из генома на протяжении тысячелетий во все более развитые виды.
Знаменитое выражение Геккеля «онтогенез обобщает филогенез» на самом деле выражает тот факт, что высшие виды повторяют на стадиях эмбрионального развития последовательность, уже обнаруженную у эволюционно предшествующих видов.
Ранние стадии эмбриона у позвоночных имеют тенденцию быть похожими, особенно до появления жабр.
У видов, которые переключаются на воздушное дыхание, затем жабры реабсорбируются и повторно используются (например, для образования эндокринных желез), но генетическая информация, относящаяся к образованию жабр, сохраняется и у людей. Это, очевидно, пример эмбриональных структурных генов, которые присутствуют в геноме всех позвоночных и должны оставаться репрессированными после того, как функционировали в их онтогенетический момент.
Интерпретация эмбриогенеза в смысле регуляции действия генов позволяет объединить сложный традиционный опыт экспериментальной эмбриологии.
ДВОЙНЯШКИ
Зигота и первые бластомеры до начала синтеза белка являются тотипотентными, то есть способны дать жизнь всему организму. С этим связаны эксперименты Спемана, который получил два эмбриона в результате удушения зиготы земноводных. Подобный феномен лежит в основе феномена однояйцевых близнецов у человека, которые именно по этой причине называются монозиготными (MZ). Подопытные близнецы Спеманна были вдвое меньше нормального размера, в то время как у человека они были совершенно нормальными. Это объясняется тем, что у амфибий два эмбриона должны были разделять единственный уже полученный желток, в то время как у человека эмбрионы могут получать через плаценту все, что необходимо для своего развития и роста.
Следует помнить, что у «мужчин две трети случаев близнецов имеют» другое происхождение: они возникают из-за случайного одновременного созревания двух фолликулов с высвобождением двух семяпочек, которые при оплодотворении дают две зиготы; В этом случае мы говорим о дизиготных близнецах (ДЗ).
Поскольку близнецы MZ, разделенные митозом из одной зиготы, имеют одинаковый геном, различия между ними должны иметь экологическое происхождение. Вместо этого геном двух близнецов DZ подобен только в той же степени, что и у любых двух братьев. Метод близнецов, широко используется в генетике человека, а также в области спорта.
У человека, у которого определенные этические соображения запрещают эксперименты, можно установить, насколько любой характер регулируется наследственными факторами: фактически, строго унаследованные признаки (например, группы крови) всегда совпадают только у близнецов MZ; что соответствие признака в MZ близко к таковому у DZ, предполагается, что факторы окружающей среды преобладают над наследственными в определении этого фенотипического признака.