В предыдущем разделе мы видели, как два регуляторных белка не позволяют головкам миозина завершить силовой удар. Только увеличение количества ионов кальция в саркоплазме позволяет высвободить эту «безопасность», установив переключатель в положение «включено». Именно присутствие кальция во внутриклеточной среде определяет начало сложных химио-механических событий, лежащих в основе сокращения мышц.
Повышение уровня кальция в саркоплазме является конечным результатом тонкого нервного контроля. Запуск сокращения происходит только тогда, когда скелетная мышца получает сигнал от двигательного нерва.
Помимо нервных структур очень важно наличие так называемого саркоплазматического ретикулума. Внутри мы находим «высокую концентрацию ионов кальция».
Саркоплазматический ретикулум
Саркоплазматический ретикулум представляет собой сетчатую канальцевую структуру, которая полностью охватывает каждое мышечное волокно, проникая во внутренние пространства между одной миофибриллой и другой. При внимательном рассмотрении можно заметить две особые структуры:
СЕТЧИКИ: они образованы продольными канальцами (которые накапливают ионы Ca2 +), которые, анастомозируя друг с другом, перетекают в более крупные трубчатые структуры, называемые терминальными цистернами, которые концентрируют и секвестрируют Ca2 +, а затем высвобождают его при появлении адекватного раздражителя.
ПОПЕРЕЧНЫЕ ТРУБОЧКИ (Т-канальцы): инвагинации клеточной мембраны (сарколемма), тесно связанные с терминальными цистернами. Покрывающая их мембрана, находясь в прямом контакте с сарколеммой, может свободно сообщаться с внеклеточной жидкостью (вне клетки).
Комплекс ПОПЕРЕЧНАЯ ТРУБКА + ТЕРМИНАЛЬНЫЕ БАКИ (размещенный по бокам) составляет так называемую ФУНКЦИОНАЛЬНУЮ ТРИАДУ.
Особая структура поперечных канальцев позволяет быстро, без задержек, передавать потенциал действия внутри мышечного волокна.
Поперечный каналец регулируется зависимым от напряжения рецепторным белком, активация которого при достижении потенциала действия стимулирует высвобождение Са2 + из концевых цистерн. Повышенная концентрация этих ионов представляет собой начальное событие сокращения мышц.

Основы сокращения мышц
Нервный импульс, исходящий из центра и переносимый мотоэнуронами, достигает уровня моторной пластинки и распространяется внутри мышечного волокна благодаря мембранной трубчатой системе. Потенциал действия и последующая деполяризация сарколеммы определяют высвобождение Са2 + из цистерн саркоплазматического ретикулума. Эти ионы, взаимодействуя с системой регуляции тропонин-тропомиозин, вызывают высвобождение активного центра на актине и, как следствие, образование актомиозиновых мостиков (см. специальную статью).
Как только стимул, вызвавший сокращение, истощается, расслабление мышц происходит посредством активного АТФ-зависимого процесса, цель которого - вернуть ионы кальция в саркоплазматический ретикулум (восстановить ингибирующий эффект системы тропонин-тропомиозин) и способствовать растворение актомиозинового мостика.
Мышечная иннервация
Сокращение мышечных волокон является результатом нервного раздражителя, который проходит через альфа-мотонейрон, пока не достигнет моторной пластинки. Тело клетки этого мотонейрона находится в вентральном роге серого вещества спинного мозга.
Несколько мышечных волокон, имеющих сходные анатомо-физиологические характеристики, иннервируются одним двигательным нейроном. Каждое из этих волокон получает афференты только от одного двигательного нейрона.
Количество волокон, контролируемых двигательным нейроном, обратно пропорционально степени тонкости и точности движения, необходимого для мышцы, которая их содержит. Экстраокулярные мышцы, например, с исключительной точностью поддерживают подвижность луковицы; по этой причине каждый мотонейрон иннервирует очень мало мышечных волокон. В других частях тела, где не требуется такой утонченности, соотношение может изменяться от 1: 5 до 1: 2000 - 1: 3000. Вообще говоря, чем меньше мышца, тем меньше двигательная единица.

Комплекс, состоящий из альфа-спинномозгового мотонейрона, его эфферентного волокна (которое выходит наружу и направляется к периферии, передающей импульс) и контролируемых мышечных волокон, составляет простейшую нейрофункциональную единицу мышцы, называемую:
НЕЙРОДВИГАТЕЛЬНЫЙ БЛОК.
Нейромоторная единица - это наименьший функциональный элемент мышцы, которым может управлять нервная система.
Вопреки тому, что можно было бы подумать, не все нервные волокна двигательной единицы направлены на соседние волокна. Фактически, мышечные волокна, принадлежащие к данной единице, смешаны с волокнами, принадлежащими другим двигательным единицам. Эта конкретная компоновка обеспечивает более широкое пространственное распределение силы, создаваемой двигательными узлами, и меньшее натяжение между пучками волокон.
Кроме того, не все нейромоторные единицы одинаковы. Они классифицируются на основе времени сокращения, пика создаваемой силы, времени релаксации и времени усталости. Это позволяет различать двигательные единицы по:
- линза типа I (или S от "Slow" или SO от "Slow Glycolitic")
- быстрый тип IIb (или FF от «Fast Fatiguing» или FG «Fast Glycolitic»)
- промежуточные продукты типа IIa (или FR от «fast fatigue resistent» или FOG «Fast Oxidative Glycolitic»).
Каждая двигательная единица состоит из мышечных волокон с однородными характеристиками. Например, все резистентные волокна относятся к медленным двигательным единицам, а к быстрым - наоборот.
Другие статьи по теме «Мышечная иннервация и саркоплазматический ретикулум»
- сокращение мышц
- мышцы человеческого тела
- Скелетные мышцы
- Классификация мышц
- Мышцы с параллельными пучками и перистыми мышцами
- Анатомия мышц и мышечные волокна
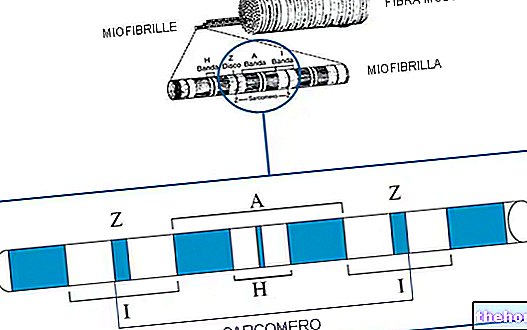
- миофибриллы и саркомеры
- актин миозин
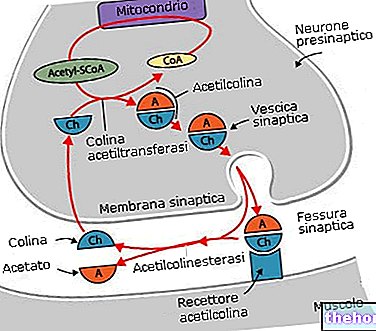
- нервно-мышечная бляшка




.jpg)