См. Также: кетогенная диета; диабетический кетоацидоз.
Общность
В прошлом считалось, что кетоновые тела являются результатом чрезмерного метаболизма, вызванного употреблением слишком большого количества жира или диабетом. Кетоновые тела, с другой стороны, естественным образом вырабатываются нашим телом: мозг приспосабливается к использованию этих метаболитов в определенных условиях. длительного голодания (у диабетиков кетоновые тела заменяют метаболизм глюкозы). Кроме того, может быть нарушение пути кетоновых тел в случае плохого питания.
Что такое кетоновые тела
Кетоновые тела являются производными липидов (они происходят из метаболизма липидов, почти исключительно печеночных), но имеют характеристики, которые делают их похожими на сахара:
- Высокая скорость ввода;
- Быстро использовать.
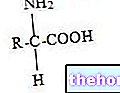
Даже некоторые аминокислоты, в частности при метаболических состояниях, могут образовывать кетоновые тела (лейцин, лизин, фенилаланин, изолейцин, триптофан и тирозин).
Биологическая роль
- Кетоновые тела имеют небольшой размер, поэтому они транспортируются очень быстро (намного больше, чем жирные кислоты, которые, с другой стороны, нуждаются в транспортных белках, таких как альбумин);
- кетоновые тела используются почти исключительно мышцами и периферическими тканями, но также и сердцем (20-30% энергии, которую оно использует, поступает из кетоновых тел) и мозгом (в случае длительного голодания).
Синтез
Кетоновые тела синтезируются ацетилкоферментом А, который образуется в результате метаболизма жирных кислот.
Фермент, катализирующий первую стадию, - это Β-кетотиолаза, который использует серу ацетилкофермента A для производства Β-кетоацил-кофермента A (это реакция, противоположная реакции, наблюдаемой в-окислении жирных кислот); эта реакция не является спонтанной, но запускается последующей реакцией , катализируемый из "гидроксиметилглутарил-кофермент А-синтаза и который включает присоединение второго ацетилкофермента A с получением 3-гидрокси-3-метилглутарил-кофермента A.
Впоследствии вмешивается литический фермент, который превращает 3-гидрокси-3-метилглутарил-кофермент А в ацетат уксуса, который является кетоновым телом. Ацетат уксуса может быть отправлен в периферические ткани или под действием фермента гидроксибутиратдегидрогеназа, превращенный в 3-Β-гидроксибутират. Если ацетат уксуса находится в очень высокой концентрации, он также может самопроизвольно декарбоксилироваться в ацетон.
Ацетон, ацетат уксуса и 3-Β-гидроксибутират - это три кетоновых тела, которые мы рассматриваем; ацетон - это отходы, которые произвольно образуются на пути кетоновых тел и удаляются при выдохе и транспирации.
Использование в периферических тканях
Кетоновые тела, вырабатываемые в печени, отправляются в периферические ткани.
Давайте посмотрим, что происходит, когда ацетат уксуса и 3-Β-гидроксибутират достигают периферических тканей. Ацетат уксуса является Β-кетокислотой, поэтому, если он активирован, его можно использовать в процессе Β-окисления для получения ацетилкофермент A: следовательно, необходимо превратить Β-кетокислоту в-кетоацил-кофермент A.
Когда ацетат уксуса попадает в митохондрию клетки периферической ткани, он подвергается действию фермента. сукцинилкофермент трансфераза: через этот фермент ацетат уксуса вступает в реакцию с сукцинилкоэнзимом А (происходящим из цикла Кребса) и образуются сукцинат и ацетилкофермент уксуса.
Используя сукцинилкофермент А для активации ацетата уксуса, мы переходим в цикл Кребса, стадию, на которой производится ГТФ: это процесс, с точки зрения энергии, который клетка готова платить, чтобы получить кофермент А ацетил уксуса. ; последний затем попадает под действие Β-кето тиолаза (Фермент Β-окисления) для производства двух молекул ацетилкофермента А, которые отправляются в цикл Кребса.
Если 3--гидроксибутират направляется в периферические ткани, последний внутри митохондрии превращается в ацетон уксуса под действием фермента Β-гидроксибутиратдегидрогеназы с образованием НАДН, что соответствует примерно 2,5 АТФ; производимый ацетат уксуса следует по ранее описанному пути.
Клетка периферической ткани потребляет больше энергии из 3-Β-гидроксибутирата, чем из ацетата уксуса, но доставка того или другого к периферическим тканям зависит от доступности энергии в печени.
C "представляет собой немаловажное количество метаболизируемых жирных кислот, содержащихся в пероксисомах, а не в митохондриях; пероксисомы представляют собой органеллы меньше митохондрий и богаты ионами металлов и ферментами пероксидазы. Ферменты пероксидазы используют перекись водорода для ускорения окислительно-восстановительных процессов, поэтому в пероксисомах есть представляет собой ферментативную систему, способную производить перекись водорода.
При-окислении в пероксисомах «ацилкофермент А получается под действием»ацилкофермент А оксидаза (В митохондриях же действовал фермент ацилкофермент А дегидрогеназа). Также в этом случае образуется транс 2,3 еноил кофермент А, который подвергается действию бифункционального фермента (он выполняет ту же функцию, что и в митохондриях с помощью «еноил-кофермента А гидратазы и L--гидроксиацил-кофермента А дегидрогеназы) и, таким образом, превращается в Β-кетоацилкофермент А. Последний, как и в митохондриях, подвергается действию Β-кетотиолазы и ацетилкофермента А. и ацилкофермент А получают с углеродным скелетом, уменьшенным на две единицы по сравнению с исходным, который возвращается в циркуляцию.