Циклическая преемственность живых существ обнаруживает связи между последовательными поколениями в явлениях воспроизводства.
Воспроизведение осуществляется на разных уровнях эволюционной шкалы, в различных ответвлениях царства растений и животных, у разных живых видов, с помощью такого разнообразия механизмов, которое само по себе оправдывает целый трактат.
Первая классификация феноменов воспроизводства должна отличать одноклеточные организмы от многоклеточных, поскольку только в первом случае деление клеток совпадает с воспроизводством.
У многоклеточных размножение может быть агамным или половым (или гамическим).
Относительно реже агамное размножение основано на механизме митоза, так что изменчивость вида скорее зависит от повторного появления мутаций.
Существуют также различные механизмы, такие как стробилизация, споруляция и т. Д., В то время как у растений мы находим регенеративные формы, хорошо известные в сельском хозяйстве (черенки, отводки и т. Д.).
Однако наиболее распространенным репродуктивным механизмом у высших форм является половой, соответствующий возникновению мейоза, образованию гамет и их слиянию в зиготе (оплодотворение).
У первобытных видов гаметы морфологически не дифференцированы: в данном случае мы говорим об изогаметии. Однако всегда есть две серии гамет, обозначенные символами (+) и (-), и оплодотворение может происходить только в результате «встречи между гаметами противоположного знака»: следовательно, существует биологическое различие, еще не выраженное морфологически.
По мере продвижения по эволюционной шкале появляется морфологическая и функциональная дифференциация, при которой тип женских гамет обычно наделен обильным резервным материалом (дейтоплазма или теленок, которые будут обеспечивать развитие эмбриона до тех пор, пока он не станет метаболически независимым), и тип мужские гаметы наделены подвижностью для достижения женских.Гаметы всегда гаплоидны и являются результатом мейоза. Их слияние дает начало диплоидной зиготе.
Между мейозом и зиготой может пройти ряд поколений гаплоидных клеток, так же как между зиготой и мейозом может пройти ряд поколений диплоидных клеток, при этом различные разновидности генеративных циклов выражаются чередованием поколений.
Вид диплонте (с диплоидным организмом) характеризуется гаметическим мейозом: мейоз непосредственно производит гаметы, которые путем слияния немедленно восстанавливают диплоидное состояние. Это распространенный случай многоклеточных животных, включая человека.
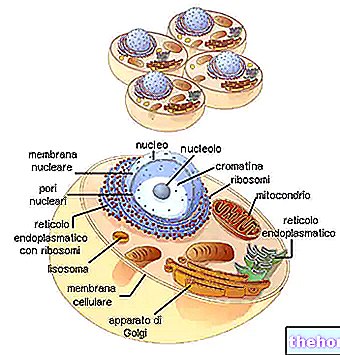
Гаметогенез
Принимая во внимание размножение гаметным мейозом, как это происходит у человека, попробуем выяснить, как мейоз вписывается в гаметогенез (образование гамет).
В эмбриональном развитии мужского и женского гаметогенеза (называемого сперматогенезом и оогенезом) происходит ранняя дифференциация между клетками, предназначенными для формирования тела (соматическая линия), и клетками, предназначенными для производства гамет (зародышевая линия). Начальные клетки зародышевой линии называются протогонами. При дифференцировке гонад в мужском или женском смысле происходит дифференцировка половых клеток соответственно на сперматогонии и овогонии.
Глядя на сперматогенез, мы видим, что в сперматогониях существует серия клеточных поколений, которая продолжается на протяжении всей жизни. Только часть непрерывно производимых сперматогоний отличается от нормального митотического цикла, и вместо него начинается мейотический.
Половая клетка, в которой начнется мейоз (редупликация, а затем первое деление), называется сперматоцитом первого порядка; его деление дает начало двум сперматоцитам второго порядка, которые при втором делении дают в общей сложности четыре сперматиды.
Мы можем градуировать редукцию хромосомного набора от 4n сперматоцитов первого порядка (после редупликации есть четыре хроматила для каждой пары гомологов) до 2n сперматоцитов второго порядка и n сперматид, как уже было показано при изучении мейоз, что, таким образом, мы заключаем. Таким образом, сперматиды уже являются гаплоидными, но они еще не являются зрелыми гаметами. Исходя из структуры клетки гаплоидного типа, функциональное созревание (называемое спермиогистогенезом) превращает сперматиды в сперматозоиды, то есть зрелые мужские гаметы.
В женском гаметогенезе (или оогенезе) есть несколько отличий. Во-первых, количество готовых гамет намного меньше. Подсчитано, что около 5 х 105 овогонов готовятся в гонадах самок человеческого вида; из них только около 400 заинтересованы в созревании фолликула и его последующем отторжении, в цикле, который обычно затрагивает только один фолликул в месяц в течение приблизительно 35 лет фертильного периода.
Различное количество гамет, приготовленных у двух полов, соответствует уже упомянутой разнице в функциях и поведении: сперматозоиды маленькие, подвижные и многочисленные в связи с необходимостью поиска яйцеклетки и низкой вероятностью ее обнаружения; большие, инертные и немногочисленные по отношению к функции обеспечения эмбриона резервным материалом и защиты, обеспечиваемой им внутренним оплодотворением (естественно, особенно при внешнем оплодотворении, яйцеклетки также должны быть более многочисленными).
Необходимость обеспечить гаметы резервным материалом соответствует наличию в «оогенезе» фазы остановки мейоза, во время которой хромосомы частично деспирализуются. Затем мы наблюдаем так называемые «хромосомы пера», в которых ряд of extroversions идентифицирует черты, при которых гены, ответственные за синтез дейтоплазмы, дерепрессированы.
Меньшее количество, необходимое для женских гамет, также соответствует тому факту, что из четырех гаплоидных клеток, продуцируемых мейозом, только одна получает весь резервный материал и становится гаметой, в то время как остальные три (полоциты или полярные тельца) содержат только хромосомный материал, не может дать начало зиготам и эмбрионам и обречен на регресс.
Удобрение
Оплодотворение, то есть встреча мужской и женской гамет, может происходить по-разному.В животном мире мы наблюдаем переход от внешнего оплодотворения (гаметы подвержены какому-либо экологическому риску и поэтому обязательно очень многочисленны у обоих полов) к внутреннему оплодотворению, с которым родительская забота также связана с метаболическими отношениями млекопитающих между матерью и плодом. .
Оплодотворение после встречи гамет противоположного пола должно происходить при соблюдении двух условий: специфичности и уникальности. То есть необходимо убедиться, что сперматозоид того же вида, что и яйцеклетка, и что, как только первый входит, другие не входят.
Специфичность обеспечивается биохимическими характеристиками акросомы и поверхности плодного яйца. Фактически, говорят о реакциях между «удобрениями» и «антифертилизинами», специфичность которых сравнима со специфичностью взаимодействия ферментов и субстрата.
Уникальность оплодотворения гарантируется изменением структуры поверхности яйцеклетки («корковая реакция»), которая начинается прямо в момент первой специфической реакции удобрения / антифертилизина; после этой реакции мембрана яйцеклетки изменяется, так что любые другие сперматозоиды, которые достигают ее, больше не могут инициировать специфическую реакцию оплодотворения.
После оплодотворения хвост сперматозоида остается внешним по отношению к яйцеклетке, а хромосомный материал проникает в него. Он, называемый «мужской пронуклеус», присоединяется к «женскому пронуклеусу» яйцеклетки, образуя диплоидное ядро зиготы.













.jpg)